
Artificial light at night (ALAN) disrupts the biological rhythms, migration routes, and reproductive cycles of nearly every wildlife group studied — from migratory songbirds to soil bacteria. European research under COST Action ES1204 (LoNNe) and the Shell L15 North Sea experiment have quantified losses that US-centric sources routinely miss. If you live near a Natura 2000 site, this is not a future problem. It is happening now, to species protected under EU law, with no lighting standards in place to stop it. For the foundational science behind what light pollution is and how it is measured, see our overview of light pollution: science, ecology, and solutions.
The Scale of the Ecological Crisis
ALAN now covers 80% of global night skies, costs ecosystems an estimated USD 3.4 trillion per year, and has affected every taxon so far examined in peer-reviewed research.

The numbers arrived in waves. Falchi et al. 2016, published in Science Advances, produced the first calibrated global atlas of artificial night sky brightness and found that more than 80% of the world’s population lives under light-polluted skies. Over 88% of Europe’s land surface qualifies. Artificial sky glow is now detectable above 22% of the world’s coastlines — environments that happen to combine high ecological sensitivity with concentrated human infrastructure. That is not a coincidence. Ports, resorts, and marinas cluster on coasts. So do sea turtles, seabirds, coastal fish, and intertidal invertebrates whose biological clocks run on genuine darkness.
The growth rate makes the static numbers worse. Kyba et al. 2017 (Science Advances) tracked Earth’s artificially lit outdoor surface between 2012 and 2016 and found it expanding at 2.2% per year in both radiance and area extent. At that rate, a landscape that is borderline light-polluted today crosses threshold values for documented ecological disruption within a planning cycle. This is not a distant problem. It is infrastructure being installed right now, at a pace that ecological science can barely track. For the full context of ALAN as a research concept and its measurement methodology, see our article on ALAN — Artificial Light at Night: the research framework.
From 22% of Coastlines to 80% of Night Skies
Skyglow — the diffuse brightening of the night sky above settled areas — is the background condition against which all other ALAN effects operate. It travels. A city’s skyglow extends measurably 200 km from its source under clear-sky conditions. For migratory species navigating at altitude, there is no local light pollution. There is only the cumulative glow of every unshielded fixture in the region beneath them, blurring the stellar backdrop they navigate by. The coastal statistic matters because coastal habitats are disproportionately rich: coral reefs, mangroves, estuaries, intertidal zones — these represent a fraction of global surface area but a vastly larger fraction of global biodiversity. Concentrating ALAN on coastlines is ecologically costly in a way that lighting equivalent land area in monoculture farmland would not be.
The USD 3.4 Trillion Ecosystem Bill
Economic valuations of ecological damage are always partial and always contested. Anderson, Kubiszewski, and Sutton 2024, published in Remote Sensing, produced one of the most methodologically careful attempts to quantify the annual loss of ecosystem service value attributable to light pollution. Their figure: USD 3.4 trillion per year — roughly 3% of global GDP, derived from comparing ecosystem service value in areas with maximum ALAN levels against dark-baseline estimates using the Copernicus PROBA-V landcover database. A 40% reduction in ecosystem service value was observed in the most heavily light-polluted areas.
The number is large enough to attract scepticism, and that scepticism is worth taking seriously. Ecosystem service valuations collapse multiple assumptions into a single figure, and the mechanisms linking ALAN to service loss — reduced pollination, altered fish yields, impaired soil function — are real but their economic translation is approximate. The number’s value is not precision. It is scale. Even if the true figure is half of Anderson et al.’s estimate, it exceeds the GDP of most European nations. That is an argument for treating ALAN as an environmental stressor with economic consequences, not merely an aesthetic problem.
How LoNNe / COST ES1204 Changed European Research
Before COST Action ES1204 — the Loss of the Night Network (LoNNe), active 2013–2017 across 27 EU member states and associated countries — European ALAN research was fragmented. Individual laboratories studied specific taxa; there was no shared measurement protocol, no interoperable dataset, no common vocabulary. LoNNe changed that. Coordinated by the Leibniz Institute of Freshwater Ecology and Inland Fisheries (IGB) in Berlin, the network produced four intercomparison field campaigns that established the European measurement baseline still in use, contributed to the development of the ALAN research field as a recognised discipline, and made the ecological and health consequences of artificial light legible to European policymakers for the first time. The research you read in this article — and the gaps it still cannot fill — are in large part shaped by what LoNNe established and what it left for subsequent researchers to pursue.
Birds and Artificial Light
Millions of birds die annually in building collisions caused by artificial light — and European offshore data show the problem has a clear, cheap, spectral solution.
Nocturnal bird migration is one of evolution’s more precise achievements. Billions of birds cross Europe each year navigating by stellar geometry, magnetic field gradients, and the polarisation of twilight. ALAN disrupts all three cues simultaneously. Attracted to lit urban cores, birds lose orientation, exhaust themselves circling light sources, and collide with illuminated glass facades. North American mortality estimates — where the monitoring infrastructure is most developed — range from 365 million to nearly one billion bird deaths per year in building collisions, with ALAN identified as a primary contributor (Loss et al. 2014, The Condor). European equivalent data are sparse, but the mechanism is identical. The Frankfurt Lights Out programme and similar European initiatives have shown measurable collision reductions when facades are darkened during peak migration windows.
The behavioural effects begin before collision. European blackbirds (Turdus merula) in lit Munich suburbs sing around one hour earlier than rural counterparts under equivalent seasonal conditions (Dominoni et al. 2013, PLOS ONE). In controlled experiments at 0.3 lux — less than the light under a street lamp at typical distance — captive blackbirds accelerated reproductive development by up to a month compared to dark-night controls. That is not merely a curiosity of urban ornithology. Breeding earlier than the food supply supports it is a mismatch that reduces reproductive success across the season. For the full species-level analysis, see light pollution and birds: migration, collision, and the Shell L15 experiment.
How City Lights Kill Migrating Birds
The mechanism is well-characterised. Migrating birds depend on stellar navigation — a celestial compass calibrated against star rotation that requires genuine darkness to read accurately. Under skyglow, the compass fails. Birds are drawn toward lit areas by the same visual hierarchy that orients hatchling sea turtles toward the ocean: they move toward the brightest available signal. In urban environments, that signal is the city skyline. Concentrations of birds above cities during migration are measurable by weather radar, and Doppler data from New York show that a single major building’s lights can redirect thousands of birds from their migration path in a single night. Glass collision is the terminal endpoint of a sequence that begins with light-induced course deviation.
Bird tetrachromatic vision — four colour receptors, including UV sensitivity — means that even wavelengths invisible to human observers contribute to navigational disruption. A facade that appears unlit to the human eye may still emit UV components that attract photosensitive migrants. This spectral dimension is underrepresented in lighting regulations, which currently specify luminance thresholds without wavelength constraints.
The North Sea Experiment: Shell L15 and the Green-Light Solution
This is the study that most collision-prevention literature ignores. In May 2007, Nederlandse Aardolie Maatschappij (NAM) — a Shell subsidiary — converted the external lighting of its offshore gas-production platform L15, situated approximately 20 kilometres off the barrier island Vlieland in the southern North Sea, from conventional mixed-spectrum sources to a green-spectrum system low in long-wavelength red light. The results, published by Poot et al. in Ecology and Society (2008), are stark: 2 to 10 times fewer migrating birds were observed circling the platform in confusion under the new lighting system compared to the old. That range — 50 to 90% fewer birds negatively impacted — was consistent across overcast nights, when the disorientation problem is worst because star navigation is unavailable.
The L15 result matters for three reasons. First, it is an industrial-scale field trial rather than a laboratory experiment — the conditions are real, the confounds are minimal, and the effect size is large. Second, the mechanism is spectral, not simply about reducing total lux: the green-light system was not dramatically dimmer, it was spectrally shifted away from the wavelengths that most strongly trigger avian attraction and disorientation. Third, the L15 platform is an offshore structure, not a city — which means the intervention is replicable at offshore wind farms, gas platforms, and lighthouses across the North Sea and Baltic, where the current spectral standard is white or red flashing lights. If 50 to 90% of bird attraction to offshore structures is preventable by a lighting specification change, the cumulative conservation gain across European offshore infrastructure is substantial.
Earlier Singing, Earlier Breeding: The Blackbird Study
Dominoni et al. 2013 documented something more insidious than collision: chronic behavioural reprogramming under low-level persistent ALAN. At 0.3 lux — a level routinely exceeded at the edge of any European town — blackbirds exposed continuously for several months advanced the onset of daily song by approximately one hour and began reproductive development weeks ahead of dark-night controls. The advancing of breeding phenology by itself might seem adaptive. It is not, when the insects that chicks depend on for food are still synchronised to the natural photoperiod. The mismatch between an ALAN-advanced breeder and a light-independent food supply is a reproductive cost invisible in the immediate season but cumulative across years and populations.
Insects Under Attack
A single streetlight can kill thousands of insects per night — and the pollination services they provide cannot be recovered by day-active pollinators alone.
Phototaxis is the instinct that pulls insects toward light sources. It is ancient, evolved in a world where the only bright nocturnal light source was the moon and bioluminescent signals. Artificial light exploits this instinct catastrophically. A single streetlamp operating through a summer night can trap and kill hundreds of insects per hour — drawn in, exhausted circling the source, killed by heat or by the predators that have learned to congregate beneath these artificial feeding stations. Across Europe, moth abundance has declined by approximately 33% since the 1970s (UK State of Nature reports), with ALAN consistently identified as a co-driver alongside habitat loss and pesticides. The insects dying at streetlights are not a uniform sample — nocturnal species and those most sensitive to short wavelengths are over-represented, which means the ecological community that returns after lamp-switch-off is already biased toward light-tolerant species.
The pollination consequence is where the individual-insect mortality scales up to ecosystem disruption. Knop et al. 2017, published in Nature, conducted a field experiment across Swiss meadows — some exposed to street-lamp-equivalent ALAN, some maintained in natural darkness. Nocturnal pollinator visits to flowering plants fell by 62% in the lit plots. Fruit set — the translation of pollination into reproductive output — dropped 13% by season’s end. Nocturnal pollination is not redundant with daytime pollination: specific plant-pollinator partnerships operate only after dark, and the Knop team showed that the loss of nocturnal visitors propagated into the daytime pollination network, which destabilised even without direct light exposure. For the full pollinator analysis, see insects at streetlights: why moths die and pollinators disappear.
Why Moths Spiral to Their Deaths
The classic image — a moth orbiting a flame — is not a death wish. The behaviour derives from the ancient navigation strategy of transverse orientation: maintaining a fixed angular relationship to a distant light source (moon, stars) to fly in a straight line. A nearby light source renders this strategy incoherent. The moth attempts to maintain its fixed angle to a light that is no longer at infinite distance, and the geometry produces an inward spiral. The moth is not attracted to the light in any motivational sense. It is doing exactly what its nervous system evolved to do — and dying because the environment has changed faster than evolution can track.
Spectral composition matters enormously here. Insects are most sensitive to ultraviolet and short-wave blue light. Cool-white LEDs at 5,000 K and above saturate the spectral range insects are most responsive to. Amber LEDs at 2,200 K dramatically reduce phototactic attraction at equivalent lumen output — a finding with direct policy implications for street lighting specification, discussed in detail in the solutions section below.
Nocturnal Pollinators and the Invisible Crisis
Twenty to thirty percent of insect-pollinated crop species — including many vegetables and fruits grown across European agricultural landscapes — are partially or wholly dependent on nocturnal pollination visits (Knop et al. 2017 supplementary data). These visits are invisible to daytime observers and absent from most standard pollinator monitoring, which operates during daylight hours. The nocturnal pollination crisis is therefore systematically underreported: the monitoring methodology that would detect it is not in routine use. Knop’s Swiss experiment remains the largest controlled field trial; its replication in other European ecotypes has been slow, and the agricultural economic loss from night-pollination collapse has not been formally quantified. This is a gap in the evidence base, not evidence of a gap in the problem.
Fireflies: A Vanishing Signal
Firefly (Photinus pyralis and related species) bioluminescent signalling operates on a tight lux threshold. Male fireflies scan for female flash responses; females respond only when the male signal is visible against a dark background. At ambient light levels above approximately 0.3 lux, male signals become indistinguishable from background light scatter, and the mate-detection system breaks down. This threshold — below the light level typical of a paved suburban street — means that firefly populations cannot persist in areas with standard outdoor lighting, regardless of habitat quality, host plant availability, or larval habitat condition. European firefly species (Lampyris noctiluca, Lamprohiza splendidula) are declining across their continental range. The proximate cause in many cases is not directly studied but ALAN is the most parsimonious explanation given the threshold data.
How ALAN Disrupts Animal Time
ALAN does not merely change behaviour — it reprograms circadian clocks at the cellular level, and the effects can be passed to offspring that have never been exposed to artificial light.
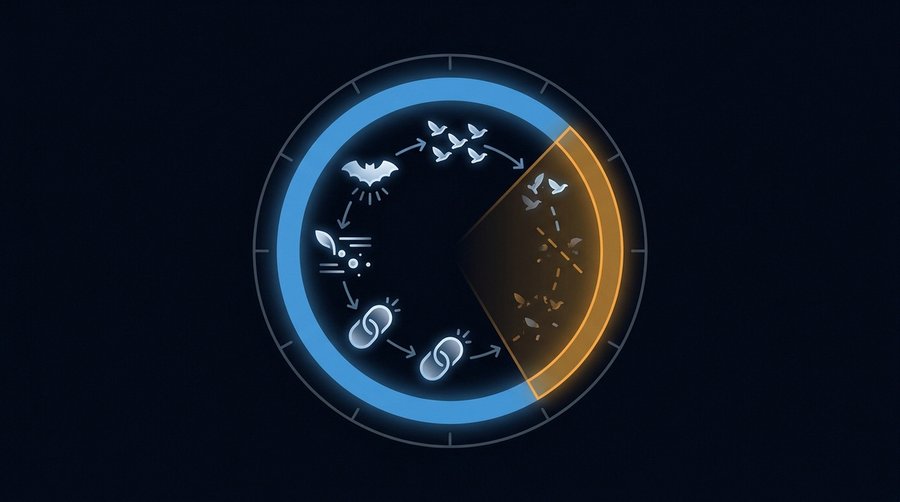
Every vertebrate runs an approximately 24-hour internal clock. Melatonin — the hormone that signals darkness to the body — is suppressed by light at intensities as low as 0.1 lux in many vertebrate species. That is roughly the illuminance on the ground beneath a streetlight at 50 metres. It is not a threshold that requires proximity to bright infrastructure. It is the background condition in the margins of most European towns. Melatonin suppression is the mechanism through which ALAN reaches circadian biology: when the pineal gland cannot secrete melatonin because retinal photoreceptors are reporting light, the downstream cascade of hormonal and metabolic cues — governing reproduction, immune function, hibernation, and migration — runs on corrupted timing. For the human parallel to this mechanism, see light pollution and human health: the science of darkness, disrupted.
The Circadian Clock in Mammals, Amphibians, and Birds
Common toads (Bufo bufo) provide one of the cleaner controlled demonstrations of ALAN’s circadian reach in amphibians. Secondi et al. exposed breeding male toads to ecologically realistic light intensities of 0.1 lux and 5 lux during the mating season. Even at 0.1 lux — the lower bound of consistent melatonin suppression in vertebrates — mating behaviour was altered and fertilisation success reduced. Males previously exposed to ALAN required multiple attempts to maintain amplexus through clutch laying, a behaviour that in undisturbed toads occurs efficiently on the first attempt. The implication is not merely behavioural: disrupted mating means reduced fertilisation rates, reduced clutch size, and reduced population recruitment across breeding seasons.
Bats present a different and particularly well-documented case. European bat species divide into light-opportunists — fast-flying aerial hawkers like Pipistrellus pipistrellus that exploit the insect concentration around streetlights — and light-avoiders — slower-flying species like Myotis daubentonii and Rhinolophus ferrumequinum that abandon foraging corridors that are illuminated. The European bat fauna of 45 species, all protected under Annex IV of the Habitats Directive, is not uniformly vulnerable to ALAN — but the most light-sensitive species are also often the most conservation-relevant, because they are restricted to specific roost types and linear landscape features that urban infrastructure routinely destroys or illuminates.
Inherited Anxiety: What Zebrafish Offspring Reveal
The most surprising result in recent ALAN research comes from zebrafish (Danio rerio). A study by researchers from the Institute of Hydrobiology of the Chinese Academy of Sciences and the Max Planck Institute of Animal Behavior, published in 2024, exposed parent zebrafish to blue-spectrum ALAN for a series of nights and then raised their offspring in complete darkness. The offspring of ALAN-exposed parents showed reduced swimming activity, reduced movement distance, and anxiety-like wall-hugging behaviour — behavioural signatures of elevated stress — despite never having been exposed to artificial light themselves. The effect was strongest at short wavelengths (365–470 nm) and was detectable in F1 offspring regardless of which parent was exposed.
This is the first documented case of transgenerational ALAN inheritance in a vertebrate. The mechanism appears to be epigenetic: ALAN exposure in the parent generation alters gene expression patterns or chromatin state in a way that is transmitted through germ cells to offspring. The conservation implication is sobering. If a population of fish, amphibians, or birds is exposed to ALAN over multiple generations — which describes most urban and peri-urban populations in Europe — the behavioural and physiological degradation does not reset when individuals move to darker habitat. The deficit propagates. Recovery may require not one generation of darkness but several.
Light Under the Midnight Sun: Nordic Seasonality
From Stockholm northward, the light environment is extreme in ways that magnify ALAN’s biological leverage. In Stockholm in June, civil twilight persists through the night — ambient light never drops below approximately 5 lux even without a single artificial fixture. For a photoperiodically sensitive species calibrated to use the last traces of twilight as a seasonal cue — to know when to begin hibernation, when to initiate gonadal development, when to synchronise migration — 1 lux of ALAN added to a 5 lux background is proportionally enormous. A 20% increase in the signal that drives annual biological timing. Reindeer (Rangifer tarandus) operate with less than 1% margin in their photoperiod detection threshold. Under typical peri-urban ALAN, that margin evaporates.
The Nordic case is not a special exception. It is an amplifier. Every ecological effect documented under ALAN conditions that are already extreme in summer is more severe, not less — because the species in these latitudes have evolved to be hypersensitive to light-dark transitions that matter. ALAN does not add noise to an already-noisy signal. It disrupts a signal that was already at the edge of biological detection.
Underwater Light Pollution
Coastal ALAN penetrates several metres underwater, shifting feeding, spawning, and algae growth across entire marine food webs — and European seas are heavily exposed.
Water attenuates light, but it does not eliminate it. Blue and green wavelengths penetrate deepest in clear coastal water — to depths of 5 to 10 metres under typical conditions, further in exceptionally transparent offshore environments. The tidal zone, estuary, and shallow reef environments that ALAN reaches most easily are exactly the environments most critical to marine food webs: spawning grounds, juvenile nurseries, and the photic zone where primary production occurs. For a detailed examination of underwater ALAN impacts, see underwater light pollution: coral reefs, salmon, and algal blooms.
Coral Reefs — 25 Nights of Light Rewire Predator-Prey Timing
Coral mass spawning is one of marine biology’s most precisely timed events. Many reef-building coral species synchronise gamete release to within hours, cued by water temperature, lunar phase, and — critically — the dimming of moonlight in the nights following a full moon. ALAN disrupts this cue. Research on reef corals in illuminated coastal environments (including Red Sea studies examining oxidative stress responses) has documented that sustained artificial illumination across the spawning window desynchronises gamete release timing and suppresses the melatonin-equivalent signals that coordinate broadcast spawning in coral polyps. Twenty-five consecutive nights of ALAN at ecologically relevant intensities are sufficient to fragment the synchrony that makes mass spawning reproductively viable. A spawning event that is desynchronised by three to four hours loses fertilisation efficiency — the gametes of one colony do not encounter those of another in the narrow window when both are viable.
Mediterranean and Adriatic coral communities are directly exposed to some of Europe’s most intensive coastal ALAN. The combination of thermal stress from warming seas and light-disruption of spawning synchrony represents a compound stressor that neither variable alone would produce.
Salmon Under Threat: Light and Predation Risk
Atlantic salmon (Salmo salar) juvenile survival in river mouths and estuaries is adversely affected by ALAN through a predation-risk mechanism. Under natural darkness, the predator-prey encounter rate between juvenile salmon and piscivorous fish that hunt by vision is low. Under ALAN at the water surface, juvenile salmon become visible to visual predators while their own schooling and evasion behaviours remain calibrated to the darkness they expect. Studies in Baltic and Norwegian river-mouth environments have documented two to three times higher predation rates on ALAN-exposed juvenile salmon compared to controls in natural darkness (Davies and Smyth 2018). This is a directly economically relevant effect — both for wild salmon populations and for aquaculture operations in semi-enclosed coastal areas where nighttime lighting of handling and maintenance facilities illuminates adjacent rearing areas.
Harmful Algal Blooms and Coastal Lighting
Photoperiod governs the growth and reproduction of phytoplankton and cyanobacteria in coastal waters. ALAN extending the effective day length for aquatic photosynthesisers selectively benefits species that are already at competitive advantage in eutrophic conditions — particularly bloom-forming cyanobacteria like Microcystis aeruginosa that produce hepatotoxins harmful to fish, waterfowl, and in severe blooms, mammals and humans. The Baltic Sea, already eutrophied by decades of agricultural nutrient runoff, is particularly susceptible to this cascade: high coastal ALAN density in the southern Baltic, combined with warm and nutrient-rich conditions, creates bloom conditions that are amplified rather than caused by ALAN but that ALAN reliably intensifies. Removing the artificial photoperiod extension that supports cyanobacterial dominance is a rarely discussed co-benefit of coastal lighting reduction.
Light Pollution as Habitat Destroyer
Light pollution fragments habitats as effectively as roads — and the solution, dark infrastructure, exists but is absent from EU policy frameworks.
Habitat fragmentation by ALAN operates differently from physical barriers. A road or fence is fixed geometry. A lit area is a volume in space-time — dark during a power outage, bright during operational hours, variably intense at different distances. But for light-averse nocturnal species, the functional effect is comparable to a physical barrier: an illuminated road, forest margin, or water body is territory they will not cross. The connectivity of the landscape at night — the ability of populations to disperse, forage, and exchange genes across the network of dark habitat — is severed by ALAN just as effectively as by fencing, and over far larger areas per fixture than a physical structure would occupy. For engineering and policy solutions to this problem, see how to reduce light pollution: engineering, policy, and ecological design and dark infrastructure: the Dutch Donkerte-Netwerk and Natura 2000 corridors.
How Light Fragments Habitats Like Roads and Fences
The habitat-fragmentation effect of ALAN is most thoroughly documented in bats. A substantial proportion of European bat species — including multiple Myotis and all Rhinolophus species — avoid commuting along lit routes even when those routes connect roost sites to foraging areas. This avoidance behaviour imposes energetic costs: bats make longer detours, arrive at foraging grounds later in the night, and accumulate less energy per foraging night. In female bats during lactation — when energy demand is at its annual peak — this loss of foraging efficiency translates directly into reduced pup survival and colony recruitment. Research on bat landscape connectivity (Rowse, Stone, Harris, and Jones — University of Bristol) has confirmed that switching from low-pressure sodium to white LED streetlighting dramatically reduces bat activity and foraging at affected routes, particularly for light-sensitive species. European hedgehogs (Erinaceus europaeus), otters (Lutra lutra), and large moths similarly treat lit road corridors as effective barriers, reducing gene flow across populations on either side of major transport arteries.
The Case for Dark Infrastructure and Biodiversity Corridors
In 2022, Sordello, Busson, and colleagues including Travis Longcore published a formal call in Landscape and Urban Planning (219: 104322) for the global development of dark infrastructure as a conservation tool. The concept is straightforward: just as green infrastructure — wildlife corridors, stepping-stone habitats, buffer zones — preserves landscape connectivity for species displaced by physical development, dark infrastructure preserves connectivity for species displaced by artificial illumination. Dark corridors along motorways, coastal zones, river banks, and forest margins — implemented through targeted lighting curfews, directional shielding, or strategic non-illumination of critical sections — can restore the movement networks that ALAN has severed.
The evidence that dark corridors work exists. In the Netherlands, an experimental stretching of unlit sections along motorway corridors showed measurable recovery of bat commuting activity within a single season. The intervention required no new infrastructure — only a decision not to illuminate specific sections that did not require illumination for human safety. The cost is near-zero. The conservation gain, in terms of restored connectivity for multiple light-sensitive taxa simultaneously, is substantial. Dark infrastructure is not an exotic conservation concept. It is the application of existing engineering decisions — what to light, how brightly, for how long — with ecological connectivity as an explicit design criterion.
Natura 2000 and the Missing Light Rules
Natura 2000 protects 18% of EU land area under the Habitats and Birds Directives. All 45 European bat species are listed under Annex IV of the Habitats Directive, affording them the highest level of legal protection: individual animals, their roosts, and their breeding sites cannot be deliberately disturbed or damaged. ALAN demonstrably disturbs roost connectivity, foraging behaviour, and reproductive success in Annex IV bat species. The logical implication — that ALAN management at and around Natura 2000 sites is a requirement of existing EU law — has not been operationalised. No Natura 2000 management plan reviewed by European conservation bodies as of the time of writing includes ALAN-specific lighting standards, lux thresholds, spectral requirements, or curfew obligations.
The EU protects the bat. It does not protect the darkness the bat needs. This is not a minor regulatory footnote. It is a structural inconsistency in EU biodiversity law that places sites designated for their highest conservation value in a legal framework that has no mechanism to address one of the most pervasive threats those sites face. The EU Biodiversity Strategy 2030 does not close this gap. Neither does the revised Habitats Directive guidance issued in the current planning period. The gap exists, is documented, and remains open.
The Forgotten Victims — Soil, Plants, and Amphibians
Below the visible ecology of birds and insects, ALAN reaches earthworms, soil bacteria, and the seasonal clock of every plant growing within range of a streetlight.

The ecological effects of ALAN that attract research attention are the dramatic ones: seabirds crashing into offshore platforms, moth populations collapsing around streetlights, coral spawn desynchronised. The effects that compound quietly across soil profiles, plant phenology, and amphibian breeding ponds are less visible, less measurable, and for that reason systematically underweighted in both the scientific literature and public awareness. They are not smaller in aggregate ecological significance. They may be larger.
Earthworms, Soil Bacteria, and Underground ALAN Effects
Earthworms (Lumbricus terrestris) emerge to the surface at night to forage and deposit organic material, forming a critical link in soil carbon cycling and structure formation. Research from IGB Berlin — including work by Maja Grubisic and colleagues — has shown that ALAN at ecologically realistic intensities of 10 lux and above significantly suppresses earthworm surface activity and foraging. In the field, this effect is compounded by the earthworm’s extreme light-aversion: even at lower intensities, surface emergence is delayed and foraging time shortened. Less foraging means less surface organic matter incorporation, slower litter decomposition, and a soil profile that processes carbon and nutrient inputs less efficiently.
A directly related finding is the ragweed link. Research published in BMC Ecology and Evolution in 2024 found that ALAN simultaneously reduces earthworm activity and increases the growth of invasive ragweed (Ambrosia artemisiifolia) — possibly because reduced earthworm activity loosens the soil competition pressure that native ground-covering plants normally exert. Ragweed is already a serious European public health problem; its flowering season extends from late summer through autumn, and its highly allergenic pollen affects tens of millions of Europeans. ALAN-facilitated ragweed expansion extends the allergenic pollen season. This is a direct pathway from a lighting specification to a public health outcome, running through soil ecology.
Plants That Forget When to Sleep
Photoperiod — the ratio of light hours to dark hours — is the master clock for plant phenology. Bud burst, flowering, leaf senescence, and winter dormancy are all cued by daylength, which in turn signals seasonal position with a precision that temperature alone cannot provide. ALAN extending the effective daylength for trees adjacent to streetlights shifts this clock. Bennie et al. replicated in Munich found that street-lit trees showed leaf-out approximately 7.9 days earlier and leaf senescence 5.4 days later than rural counterparts — an effective lengthening of the growing season by almost two weeks. In isolation this might appear benign. In context it is a phenological decoupling: an extended plant growing season that is no longer synchronised with the insect emergence, bird migration, and soil organism activity calendars that co-evolved with it. Autumn leaf fall that arrives nearly two weeks late means a compressed window for decomposer organisms to process litter before winter. Bud burst two weeks early means exposure to late frosts that a naturally-timed plant would have avoided.
Frogs and Toads: Disrupted Breeding in Urban Wetlands
Amphibians use both temperature and photoperiod to time their breeding migrations, and ALAN disrupts the photoperiod component. Common toad populations in urban and peri-urban wetlands across central Europe — including long-monitored sites in the Berlin and Stockholm regions — have declined by 40 to 60% over the past two decades, with ALAN consistently identified as one of several co-drivers alongside road mortality, habitat loss, and disease. The breeding disruption mechanism is dual: altered cue timing leads to earlier or later pond arrival, and even when the pond is reached, low-level ALAN during the mating period (at the 0.1 lux threshold documented by Secondi et al.) reduces mating efficiency and fertilisation success. Wetlands within 200 metres of streetlighting — which describes the majority of European urban ponds — operate under chronically disrupted photoperiod conditions for the amphibians that depend on them.
What Works — Evidence-Based Solutions
Turning lights off, switching to amber, and shielding fixtures demonstrably reduces ALAN impacts — the evidence is clear and the costs are low relative to the ecological return.
The technical solutions to ecological light pollution are understood. The implementation gap is regulatory and economic, not scientific. Several European programmes have demonstrated that meaningful reductions in ALAN impact are achievable within a single planning cycle when the will is present. The evidence base for what works — and at what scale — is now large enough to move beyond pilot programmes to standard practice.
Lights Out Programmes: From Houston to Helgoland
Lights Out initiatives — switching off or dimming non-essential facade, advertising, and interior lighting during peak migration windows — have produced the strongest and most replicable evidence for rapid ALAN impact reduction. The Chicago Lights Out programme, operating since 2021, has documented a 60% reduction in bird-building collisions during spring and autumn migration periods in participating buildings. The Helgoland offshore island programme — in the southern North Sea, within 50 kilometres of the Shell L15 trial site — has demonstrated equivalent reductions in seabird mortality at the island’s lighthouse and associated infrastructure when spectral and directional lighting modifications are applied. These are not marginal effects. A 60% reduction in collision mortality, applied across even a fraction of Europe’s illuminated building stock during the eight to ten weeks of peak migration, represents millions of individual birds per year.
The Frankfurt, Brussels, and Paris curfew experiments have shown that switching off decorative and advertising lighting between midnight and 5 a.m. reduces sky-glow contribution from urban centres by 15 to 25% at measurement sites outside the city — because the lighting categories switched off are disproportionately upward-directed, high-intensity, and poor-spectral-quality. The energy savings from these curfews cover the administration cost within the first year.
Amber Over Blue: Spectral Choices That Protect Wildlife
Spectral composition is the most consequential single variable in wildlife-protective lighting design. Insects are maximally attracted to UV and short-wave blue light — the spectral peak of cool-white LEDs at 4,000 to 6,500 K. Amber LEDs at 2,200 to 2,400 K reduce phototactic insect attraction by a factor of five to ten at equivalent lumen output. For bats, the evidence is direct: a controlled Dutch motorway trial that replaced white LED streetlighting with amber alternatives documented a 54% reduction in the barrier effect for light-averse Pipistrellus species along the corridor. The bats resumed commuting through the previously avoided section within weeks of the spectral switch.
Below 0.1 lux — the lower threshold of consistent melatonin suppression across vertebrate taxa — most documented ALAN effects disappear or become statistically undetectable. This means that combining spectral shift (amber), directional shielding (full cut-off fixtures), and intensity reduction (to below 0.1 lux in ecological margin zones) is a package that, where implemented, eliminates the majority of measured ALAN impacts for multiple taxonomic groups simultaneously. These are not exotic engineering specifications. They are available off-the-shelf from standard lighting suppliers and are less expensive per lumen than the blue-rich alternatives that currently dominate public procurement.
SQM Monitoring in Protected Areas as a Citizen Science Tool
Effective ALAN management requires measurement — knowing what the baseline sky brightness is in a protected area before any intervention, and tracking change over time. The Sky Quality Meter (SQM), an instrument that reports sky darkness in astronomical magnitudes per square arcsecond, has become the standard low-cost instrument for this purpose. SQM readings from Natura 2000 sites are now included in peer-reviewed European biodiversity assessments — including assessments that link quantified ALAN levels to bat activity, amphibian breeding success, and insect community composition. The instrument costs around EUR 100 and is operable by any motivated citizen observer. For more on SQM variants, monitoring methodology, and how to contribute data to research-grade networks, see measuring light pollution: methods, data, and research tools and our SQM buyer’s guide.
COST Action ES1204 (LoNNe) ran four intercomparison campaigns specifically assessing SQM reliability in field conditions and established the SQM as a valid research-grade instrument when used with consistent protocol. The result is a continent-wide citizen science infrastructure for ALAN monitoring that costs almost nothing to maintain and produces data quality sufficient for peer-reviewed publication. European protected-area managers who are not yet SQM-monitoring their sites are leaving a measurable, low-cost evidence base on the table.
Dark Sky Certification and Natura 2000: A Path Forward
International Dark-Sky Association (IDA) certification — a process that requires documented sky-quality baseline, a lighting management plan, and ongoing monitoring — provides the institutional framework that ALAN management in protected areas currently lacks in EU law. Westhavelland Dark Sky Reserve in Germany, certified in 2014 and located 60 kilometres from Berlin, has documented measurable reduction in light-averse species displacement since certification, corroborated by SQM monitoring data. Brecon Beacons in Wales has shown equivalent improvements in bat activity along buffer-zone corridors following implementation of its lighting management plan.
The logical integration — requiring ALAN assessment and management plans as a component of Natura 2000 site management plans under the existing framework of the Habitats Directive — requires no new EU legislation. It requires only that existing Directive obligations regarding species disturbance be interpreted consistently with the documented evidence on how ALAN disturbs those species. That interpretation has not happened at the level of European Commission guidance. It should. For more on dark sky places and certified refugia across Europe, see dark sky places in Europe: parks, reserves, and the science of protected night skies.
Frequently Asked Questions
Does turning off lights really help wildlife recover immediately?
For some species, recovery is measurable within days. Bird collision rates at buildings participating in Lights Out programmes drop 60% or more in the first migration season. Insect activity around de-lit streetlights recovers within hours of lamp-off. Long-term population recovery — for amphibians, bats, and earthworm communities — takes years or decades, because population dynamics operate on generational timescales. But recovery begins with ALAN removal, and the evidence from dark sky reserves like Westhavelland and Brecon shows that measurable wildlife recovery is achievable within single management plan periods when light pollution is actively managed rather than simply designated as undesirable.
What type of light is least harmful to nocturnal animals?
Amber light at 2,200 to 2,400 K, combined with full cut-off shielding that directs light downward only, and intensity kept below 0.1 lux at ecologically sensitive margins. UV and cool-white light at 4,000 K and above is maximally disruptive across the widest range of taxa. Direction matters as much as spectrum: a correctly shielded amber fixture produces less ecological impact than an unshielded amber fixture at the same lumen output, because the shielded version does not project light into the sky or horizontally into adjacent habitat. Motion-activated controls that reduce output to zero when no human is present are the most effective single addition to any ecologically sensitive outdoor lighting specification.
How does light pollution interact with other biodiversity stressors like habitat loss?
ALAN acts as a multiplier for existing stressors, not an independent parallel pressure. Anderson et al. 2024 found that populations already under stress from habitat loss, pesticide exposure, or climate disruption are two to three times more vulnerable to ALAN than populations in otherwise intact habitat. The mechanism is resilience depletion: a population that is already operating near its reproductive minimum has no buffer for additional chronobiological disruption. Removing ALAN from an otherwise degraded habitat does not restore the habitat. But adding ALAN to an already-stressed population reliably accelerates decline, and removing it reliably reduces one pressure in the compound stressor profile.
Can dark sky reserves actually protect biodiversity?
Yes — if they are managed with quantified ALAN monitoring and active lighting controls, not merely labelled. Certification without management produces no measurable ecological benefit. Westhavelland in Germany and Brecon Beacons in Wales are the two European sites with the longest documented post-certification monitoring records, and both show measurable improvements in light-sensitive species indicators within the reserve boundaries and, to a lesser degree, in buffer zones. The critical element is a sky-quality baseline established before certification, against which subsequent measurements can be compared. A Sky Quality Meter network in and around the reserve provides this for a total instrument cost that is negligible against any conservation budget.
Why are European coastlines particularly vulnerable to ALAN?
Two factors converge. First, European coasts concentrate the highest-intensity ALAN sources: resort lighting, harbour and marina installations, aquaculture facilities, and offshore oil and gas platforms. Second, coastal habitats have disproportionate ecological sensitivity: sea turtle nesting beaches in the Mediterranean, seabird colonies across the North Sea and Atlantic margins, coral and gorgonian communities in the Adriatic and Tyrrhenian seas, and juvenile salmon estuaries in the Baltic and Norwegian fjords. Twenty-two percent of the world’s coastlines are already under measurable artificial sky glow, and coastal human populations are projected to more than double by 2060. The intersection of maximum ALAN growth trajectory and maximum ecological sensitivity is, geographically, the European coastline.